Research
Vertebrate Eye Development |
|
|
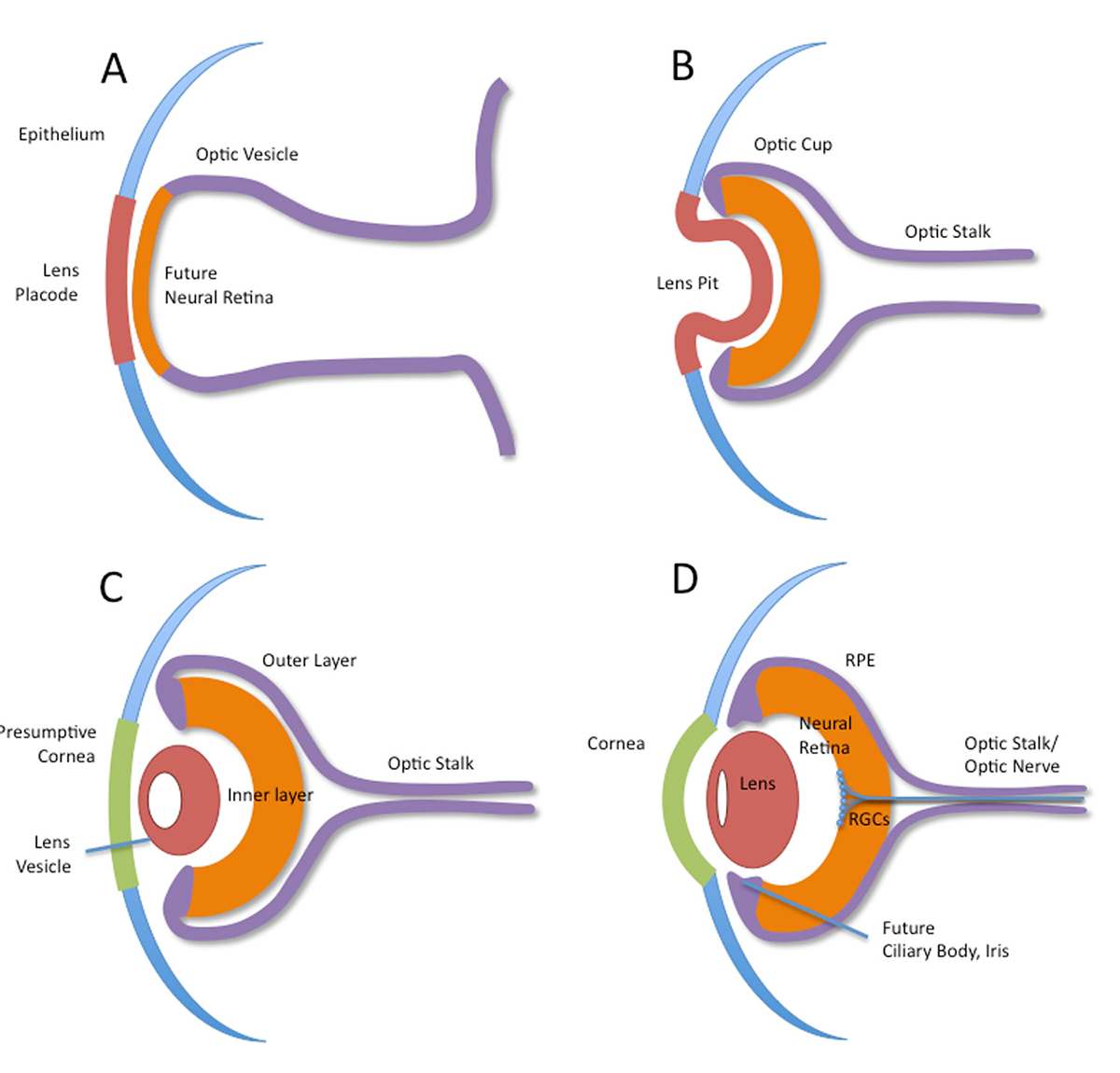
What Controls Optic Vesicle Outgrowth and Patterning? |
|
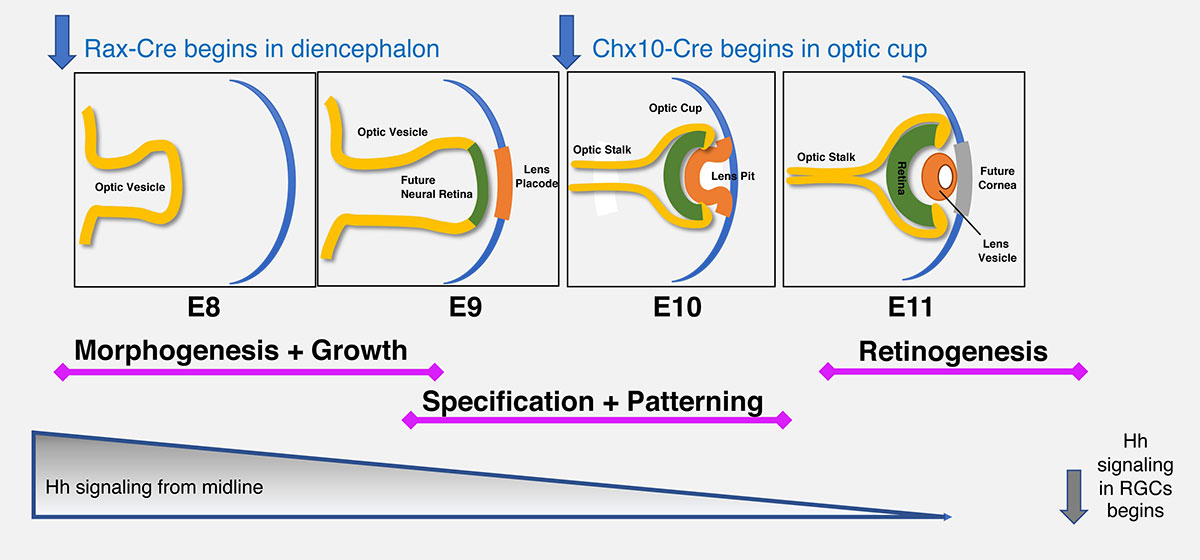
Another commonly used signaling system, the Notch pathway, relies on physical interactions between membrane-bound ligands and receptors on adjacent cells, severely limiting its range of action. During retinal development, this pathway orchestrates the appearance of distinct neuronal and glial cell types. Notch ligand-receptor binding induces sequential proteolytic cleavages to the receptor protein, ultimately releasing the intracellular domain (N-ICD) to form a nuclear protein complex with Rbpj, MAML, p300 and other proteins. Then the complex transcriptionally activates downstream genes, for example Hes1. Notch pathway mutations are linked to human birth defects such as Alagille Syndrome and Tetralogy of Fallot, SIgnal dysregulation can cause multiple types of cancer. We are exploring genetic and molecular intersections between the Shh and Notch signaling pathways during ocular growth, morphogenesis and patterning. We are using two spatiotemporally distinct Cre drivers (Rax-Cre and Chx10-Cre) to examine the phenotypes caused by loss- or gain-of-function of each pathway. |
Vertebrate Retinal Neurogenesis |
|
|
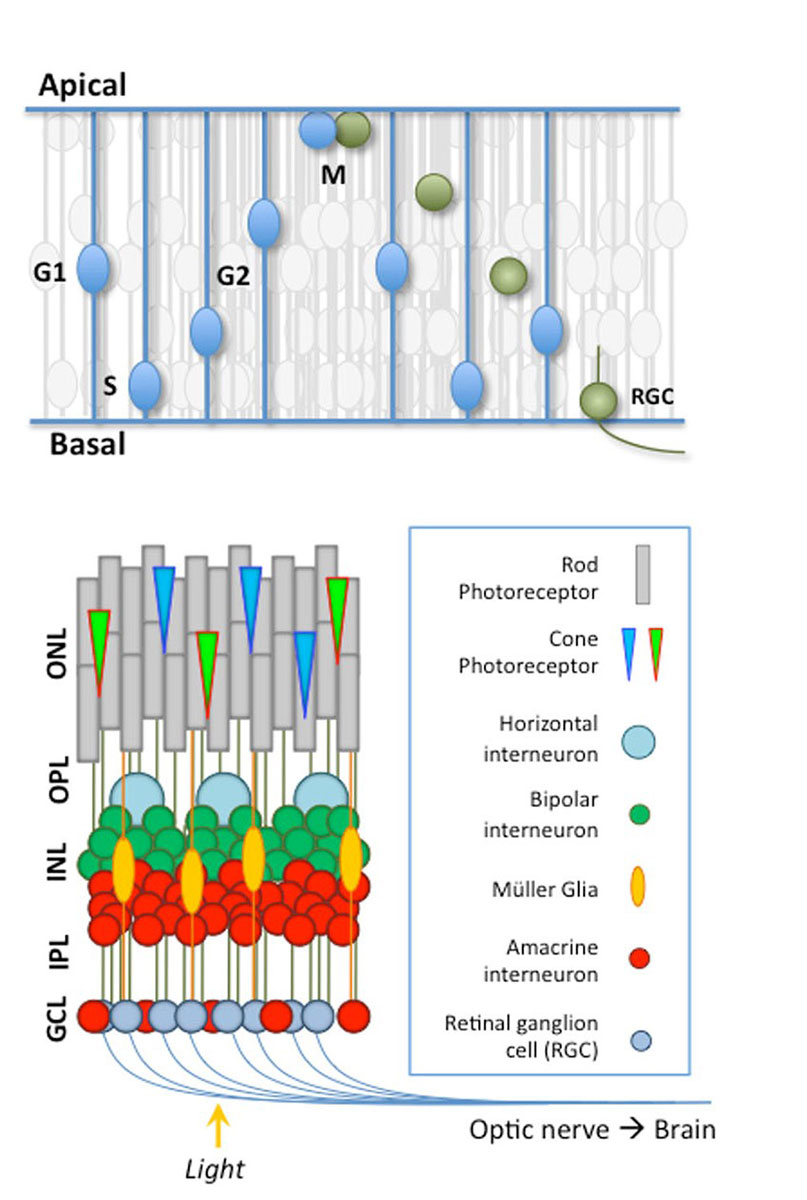
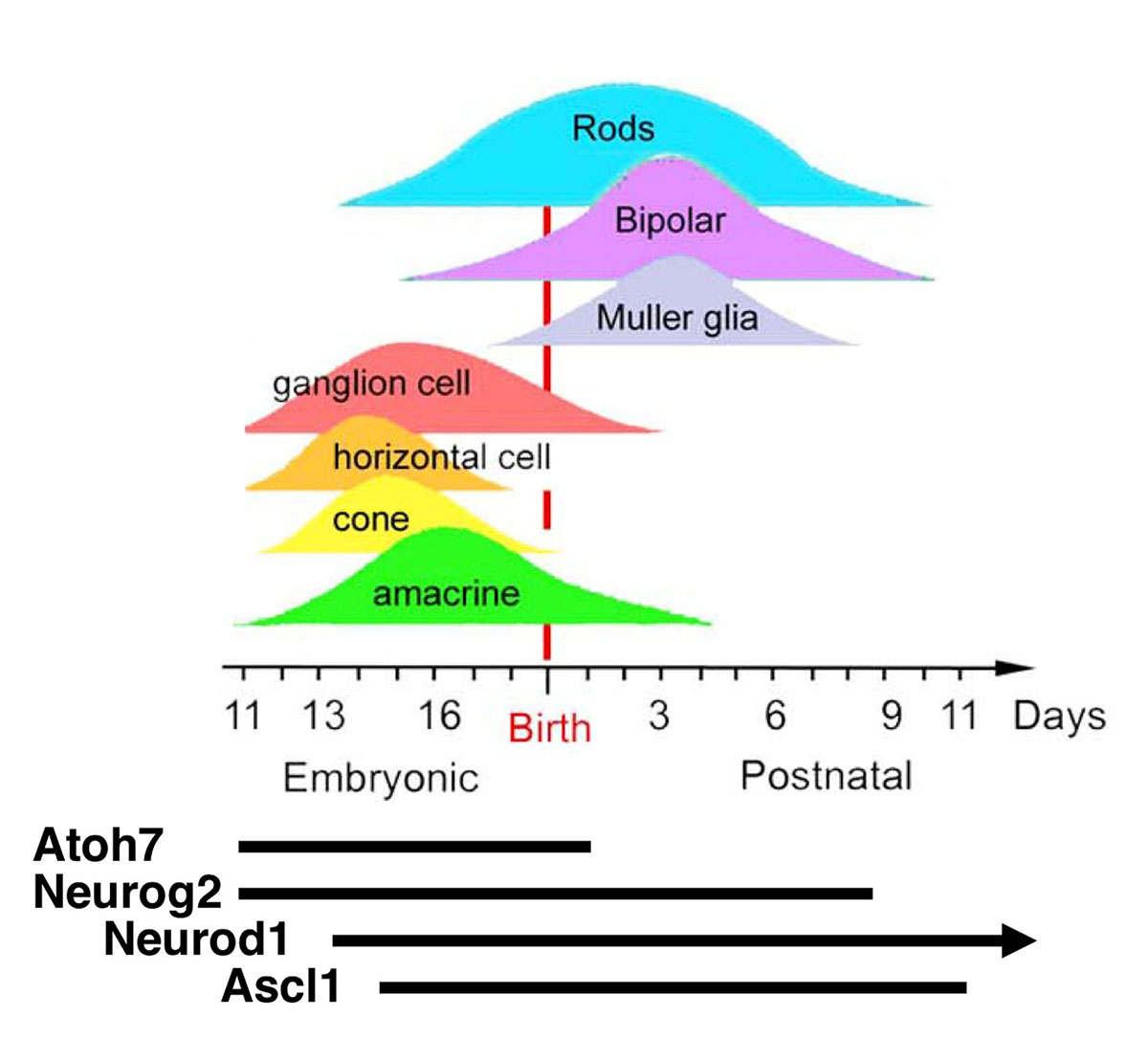
bHLH Transcription Factors in the Retina |
|
|